1. История открытия и названия нуклеиновых кислот
3. Получение нуклеиновых кислот
4. Химические свойства нуклеиновых кислот
5. Применение нуклеиновых кислот
6. Занимательные факты о нуклеиновых кислотах
1. История открытия нуклеинов и их названия
Открытие нуклеиновых кислот связано с именем молодого врача из города Базеля (Швейцария) Фридриха Мишера . После окончания медицинского факультета Мишер был послан для усовершенствования и работы над диссертацией в Тюбинген (Германия) в физиолого-химическую лабораторию, возглавляемую Ф. Гоппе-Зейлером . Тюбингенская лаборатория в то время была известна ученому миру. Пройдя практику по органической химии, Мишер приступил к работе в биохимической лаборатории. Ему было поручено заняться изучением химического состава гноя. Молодой ученый не возражал против предложенной темы, так как считал лейкоциты, присутствующие в гное, одними из самых простых клеток.
Путём многочисленных опытов он получил из гнойных клеток вещество ядерного происхождения. Мишер был уверен именно в ядерном его источнике. Поэтому он начал более тщательное выделение ядер. В то время еще никто в биохимических лабораториях не пытался выделить ядра или какие-либо другие субклеточные компоненты, так что и здесь он был пионером.
Продолжив дальше очищать ядро от других клеточных фрагментов, он получил странное вещетво. Оно не разлагалось протеолитическими ферментами, значит, не являлось белком. Отсутствие растворимости в горячем спирте указывало на то, что это вещество не являлось и фосфолипидом. По-видимому, оно относилось к новому классу биохимических соединений.
Но Мишер с большой горячностью настаивал на точности своих результатов и добивался разрешения опубликовать их в печати. Тогда Гоппе-Зейлер решил проверить данные Мишера лично. Он и два его ассистента (одним из них был русский химик Любавин) в течение года шаг за шагом прошли все этапы аналитической работы Мишера и полностью подтвердили его данные, выделив нуклеин из клеток крови и из дрожжей.
В 1871 г. работа Мишера вместе с подтверждающими ее контрольными работами Гоппе-Зейлера и его ассистентов увидела свет. Существование нуклеина как специфического ядерного вещества стало научным фактом . Вскоре методика Мишера была применена для выделения нуклеина из различных тканей.
Термин «нуклеиновые кислоты» был предложен в 1889 : нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами - из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота.
2. Нахождение нуклеиновых кислот в природе
Нуклеиновые кислоты в природе встречаются во всех живых клетках. Живые клетки, за исключением сперматозоидов, в норме содержат значительно больше рибонуклеиновой, чем дезоксирибонуклеиновой кислоты. На методы выделения дезоксирибонуклеиновых кислот оказало большое влияние то обстоятельство, что, тогда как рибонуклеопротеиды и рибонуклеиновые кислоты растворимы в разбавленном (0,15 М) растворе хлористого натрия, дезоксирибонуклеопротеидные комплексы фактически в нем нерастворимы.
Поэтому гомогенизированный орган или организм тщательно промывают разбавленным солевым раствором, из остатка с помощью крепкого солевого раствора экстрагируют дезоксирибонуклеиновую кислоту, которую осаждают затем добавлением этанола.
В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеотид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
3. Получение нуклеиновых кислот
В клетках нуклеиновые кислоты связаны с белками, образуя нуклеопротеиды. Выделение нуклеиновых кислот сводится к очистке их от белков. Для этого препараты, содержащие нуклеиновые кислоты, обрабатывают ПАВ и экстрагируют белки фенолом. Послед, очистка и фракционирование нуклеиновых кислот проводятся с помощью ультрацентрифугирования, различных видов жидкостной хроматографии и гель - электрофореза. Для получения индивидуальных нуклеиновых кислот обычно используют различные варианты последнего метода.
Современные методы химического синтеза нуклеиновых кислот позволяют получать крупные фрагменты ДНК, в том числе целые гены. Методические основы химически - ферментативных методов синтеза ДНК разработаны X. Кораной.
Они включают:
Химический синтез комплементарных, взаимоперекрывающихся олигонуклеотидов, из которых затем в результате комплементационных взаимодействий выстраиваются дуплексы - фрагменты молекулы синтезируемой ДНК с несовпадающими разрывами в обеих цепях;
Соединение (лигирование) таких олигонуклеотидов в составе дуплекса с помощью фермента Т4 ДНК-лигазы. Сборку протяженных ДНК из синтетически однотяжевых олигонуклеотидов проводят в несколько этапов. Сначала собирают небольшие дуплексы с "липкими" концами (однотяжевыми комплементарными участками), из которых затем последовательно формируют более протяженные структуры. Таким образом могут быть получены искусственные фрагменты ДНК большой длины и с любой нуклеотидной последовательностью. С помощью генетической инженерии возможно клонирование (получение в индивидуальном виде и размножение) искусственных ДНК.
Несмотря на малую эффективность этого метода, были синтезированы олигонуклеотиды, содержащие до 16 звеньев, из которых были собраны первые синтетические гены. Фосфодиэфирный метод образования межнуклеотидных связей, использованный Кораной, имеет историческое значение. Однако разработанные им приемы введения и избирательные удаления защитных групп широко используются в других методах синтеза нуклеиновых кислот.
Важным шагом в совершенствовании синтеза олигонуклеотидов явилась разработка так называемого фосфотриэфирного метода . Образующийся динуклеотид после частичного деблокирования фосфата конденсируют аналогичным образом с другими динуклеотидом и т.д. Применение этого способа, в котором используют защиту фосфатной группы, позволило значительно сократить время синтеза и повысить выходы олигонуклеотидов.
Параллельно этим методам, которые осуществляют в растворах, разрабатывались твердофазные способы синтеза нуклеиновых кислот. В последнем случае процесс проводят в двухфазной системе; нуклеозидный компонент связан ковалентно с нерастворимым полимером, а нуклеотидный компонент и необходимые реагенты находятся в растворе.
Обычно в этом случае на первой стадии нуклеозид присоединяют с помощью "якорной" группы к нерастворимому полимеру. Затем его 5"-гидроксильную группу деблокируют и конденсируют с нуклеотидным компонентом. У образующегося полностью защищенного динуклеозидмонофосфата деблокируют защитную группу в положении 5" и присоединяют следующему нуклеотид и т.д.
Наиболее распространенные методы твердофазного синтеза олигонуклеотидов основаны на использовании нуклеотидного компонента, содержащего Р( III ). В так называемом амидофосфитном способе нуклеотидным компонентом является эфир 3"-амидофосфита дезоксинуклеозида. Достаточно устойчивые амидофосфиты при протонировании в присутствии тетразола превращаются в сильные фосфорилирующие агенты. После завершения синтеза удаляют защитные группы с межнуклеотидных фосфатов, отделяют олигонуклеотид от носителя, деблокируют группы NH2 гетероциклов. Липофильную группу (МеО)2Тr удаляют после первого хроматографического разделения.
Стандартность операций в твердофазном синтезе олигонуклеотидов явилась основой для автоматизации процесса . Принцип работы автомата-синтезатора основан на подаче в реактор с помощью насоса (под контролем микропроцессора) защищенных нуклеотидных компонентов реагентов и растворителей по заданной программе в колонку, содержащую полимерный носитель с закрепленным на нем первым нуклеозидом. После окончания синтеза и отделения полностью защищенного олигонуклеотида от полимерного носителя проводят деблокирование, очистку и анализ синтезированных фрагментов ДНК. Так, с помощью гидрофосфорильного метода в автомате - синтезаторе за несколько часов получают 30-40-звенные олигонуклеотиды; возможен синтез более чем 100-звенных фрагментов ДНК. Разработаны синтезаторы, позволяющие проводить одновременно синтез несколько олигонуклеотидов.
Синтез олигорибонуклеотидов ферментативным путем осуществляют обычно с использованием рибонуклеаз или полинуклеотидфосфорилаз.
В качестве нуклеотидного и нуклеозидного компонента применяют мономеры или олигонуклеотиды. Эту реакцию используют для синтеза ди-, три- и тетрарибонуклеотидов. При увеличении длины олигорибонуклеотида начинает преобладать обратная реакция (гидролиз олигонуклеотида).
Химический синтез олигорибонуклеотидов проводят в основном с использованием тех же приемов, как и при синтезе ДНК.
4. Химические свойства нуклеиновых кислот
Нуклеиновые кислоты :
Хорошо растворимы в воде
Практически не растворимы в органических растворителях.
Очень чувствительны к действию температуры и критических значений уровня pH.
Нуклеиновые кислоты содержат в себе генетический материал всех живых организмов. Выяснение их структуры открыло новую эру в наших знаниях о природе.
Составными частями нуклеиновых кислот являются нуклеотиды. Молекула нуклеотида состоит из пентозы, азотистого основания и фосфорной кислоты. В зависимости от типа сахара различают рибонуклеиновую кислоту ( РНК ; в её состав входит рибоза) и дезоксирибонуклеиновая кислота ( ДНК ; в её состав входит сахар дезоксирибоза, у которого на один атом кислорода меньше). В обоих типах нуклеиновых кислот содержатся четыре типа оснований: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т; в РНК вместо него содержится урацил (У)). Первые два основания относятся к классу пуринов , остальные – к пиримидинам . Фосфорная кислота определяет кислотные свойства нуклеиновых кислот.
Соединяясь друг с другом фосфодиэфирной связью (3"-фосфатная группа одного и 5"-сахар другого нуклеотида), два нуклеотида образуют динуклеотид. При синтезе полинуклеотидов этот процесс повторяется миллионы раз. Фосфодиэфирный мостик является прочной ковалентной связью, обеспечивая всей цепи стабильность и уменьшая риск «поломок» ДНК.
Выяснить структуру ДНК удалось в 1953 году английским ученым Д. Уотсону и Ф. Крику . Они показали, что ДНК состоит из двух полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе цепи свиты вместе, образуя двойную спираль. Шаг спирали составляет 3,4 нм (по 10 пар оснований в витке), а диаметр витка – 2 нм. Фосфатные группировки находятся снаружи спирали, а азотистые основания – внутри. ДНК – очень хрупкая молекула, простое перемешивание её раствора может привести к разрыву цепей на более мелкие куски.
Число адениновых оснований в любой ДНК равно числу тиминовых оснований, число гуаниновых оснований всегда равно числу цитозиновых оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но эта последовательность в одной цепи полностью определяет собой последовательность нуклеотидов в другой. Пары соединяются водородными связями между основаниями в строго определённом порядке (аденин с тимином, гуанин с цитозином). Таким образом, цепи двойной спирали комплементарны друг другу.
Для того, чтобы ДНК являлась генетическим материалом, она должна быть способна нести в себе закодированную информацию и точно воспроизводиться ( реплицироваться ). Последующие исследования доказали, что ДНК действительно содержит в себе генетическую информацию.
Молекула РНК в отличие от ДНК состоит, как правило, из одной цепи и имеет гораздо меньшие размеры. Существует три основных вида РНК: транспортная (т-РНК), информационная (и-РНК) и рибосомная (р-РНК). Информационная РНК (и-РНК) является матрицей, которую рибосомы используют при синтезе белка . Её нуклеотидная последовательность комплементарна сообщению, содержащемуся в определённом участке ДНК. Транспортные РНК переносит аминокислоты к месту синтеза. Несколько видов р-РНК являются основным компонентом рибосом . Нуклеотидные последовательности т-РНК и р-РНК также определяются определёнными участками ДНК.
ДНК находится, главным образом, в ядре клетки (у прокариот рассредоточена по клетке), являясь основным веществом хромосом. РНК сконцентрирована в ядрышке, цитоплазме и частично в хромосомах. Молекул РНК в клетке значительно больше (иногда их десятки тысяч), чем молекул ДНК.
Роль нуклеотидов заключается не только в синтезе нуклеиновых кислот. Некоторые нуклеотиды играют важную роль в жизнедеятельности организмов, являясь коферментами . Примером могут служить аденозинфосфорные кислоты , содержащие аденин, рибозу и несколько остатков фосфорной кислоты. Присоединение каждой новой фосфатной группы к кислоте сопровождается аккумуляцией энергии, а их отщепление – выделением. Превращение аденозинтрифосфорной кислоты (АТФ) в аденозиндифосфорную (АДФ) является основой энергетического обмена внутри клетки.
Нуклеиновые кислоты выполняют в организме человека следующие функции: ДНК - хранение наследственной информации, РНК - реализацию генетической информации и каталитическую функцию.
Строение нуклеиновых кислот
Нуклеиновые кислоты представляют линейные полимеры нуклео- зидмонофосфатов, то есть полинуклеотиды. Нуклеотиды построены из трех компонентов: пиримидинового или пуринового основания, углевода (пентозы) и фосфорной кислоты. Нуклеотиды связаны между собой в цепь фосфодиэфирной связью. Она образуется за счет этерификации ОН-группы С-З-пентозы одного нуклеотида и ОН-группы фосфатного остатка другого нуклеотида.
Молекула нуклеиновой кислоты имеет два уровня структурной организации.
Первичная структура нуклеиновых кислот определяется как последовательность нуклеотидных остатков в полимерной цепи. Многообразие молекул ДНК и РНК объясняется их первичной структурой.
Вторичная структура у молекул ДНК и РНК разная. Молекула ДНК представляет собой правозакрученную спираль, состоящую из двух полинуклеотидных цепей с антипараллельным ходом. Это означает, что 3-концу одной цепи соответствует 5-конец другой цепи, и наоборот. Остатки оснований направлены внутрь спирали. На один виток спирали приходится 10 пар оснований. Цепи ДНК не идентичны, так как нуклеотидный состав их различен, однако первичная структура одной цепи предопределяет нуклеотидную последовательность другой цепи, то есть они комплементарны друг другу. Это связано с существованием комплементарных пар оснований. Физико-химическую основу комплементарности составляют водородные связи, которые могут образоваться только между аденином одной цепи и тимином другой, противоположно направленной цепи (пара А-Т), и аналогично между гуанином и цитозином (пара Г-Ц).
Молекула РНК состоит из одной полинуклеотидной цепи. Отдельные участки этой цепи (до 20-30 нуклеотидных пар) могут быть комплементарны между собой и образуют спиральную структуру за счет связей между аденином и урацилом (пара А-У) и гуанином и цитозином (пара Г-Ц). Между спирализованными участками располагаются одноцепочечные петли. Существует несколько разновидностей РНК: матричная (мРНК), транспортная (тРНК), рибосомная (рРНК).
Классификация нуклеиновых кислот
В одну молекулу нуклеиновых кислот может входить углевод только одного вида - рибоза или дезоксирибоза. На этом основании все нуклеиновые кислоты делятся на два типа: рибонуклеиновые - РНК (содержат рибозу) - и дезоксирибонуклеиновые - ДНК (содержат дезоксирибозу).
Физико-химические свойства нуклеиновых кислот
1. Денатурация.
Вторичная структура ОЫА стабилизируется лишь слабыми водородными и гидрофобными связями, следовательно, ОЫА способна к денатурации (плавлению) при повышении температуры до 80-90 °С. При денатурации двухспиральная молекула ДНК разделяется на отдельные цепи. Температура, при которой 50 % ДНК денатурировано, называется температурой плавления и зависит от качественного состава ДНК.
2. Ренатурация.
Если раствор денатурированной ДНК медленно охлаждать (отжиг), то вновь возникают слабые связи между комплементарными цепями, и может получиться спиральная структура, идентичная исходной (нативной).
3. Растворимость в воде.
В воде ДНК образует вязкие растворы, при нагревании таких растворов до 60 °С или при действии щелочей двойная спираль распадается на две составляющие цепи.
4. Молекулярная масса.
Молекулярная масса нуклеиновых кислот сильно варьирует, но в целом очень большая, особенно у ДНК. В ядре клетки человеческого организма содержится 46 молекул ДНК, в составе каждой из них - 3,5 млрд пар мононуклеотидов. В митохондриях есть циклическая ДНК, ее молекула содержит 16 тыс. пар мононуклеотидов. Сначала была расшифрована структура митохондриальной ДНК. В ней закодирована информация о строении 13-ти полипептидных цепей, 2-х рибосомальных РНК и 22-х транспортных РНК.
Существует два типа нуклеиновых кислот - дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза - в ДНК, рибоза - в РНК) и остаток фосфорной кислоты.
В ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, - аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований - аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований
Молекулы ДНК и РНК существенно различаются по своему строению и выполняемым функциям.
Молекула ДНК может включать огромное количество нуклеотидов - от нескольких тысяч до сотен миллионов (поистине гигантские молекулы ДНК удается «увидеть» с помощью электронного микроскопа). В структурном отношении она представляет собой двойную спираль из полинуклеотидных цепей , соединенных с помощью водородных связей между азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи прочно удерживаются одна возле другой.
При исследовании различных ДНК (у разных видов организмов) было установлено, что аденин одной цепи может связываться лишь с тимином, а гуанин - только с цитозином другой. Следовательно, порядок расположения нуклеотидов в одной цепи строго соответствует порядку их расположения в другой. Этот феномен получил название комплементарности (т. е. дополнения), а противоположные полинуклеотидные цепи называются комплементарными. Именно этим обусловлено уникальное среди всех неорганических и органических веществ свойство ДНК - способность к самовоспроизведению или удвоению . При этом сначала комплементарные цепи молекул ДНК расходятся (под воздействием специального фермента происходит разрушение связей между комплементарными нуклеотидами двух цепей). Затем на каждой цепи начинается синтез новой («недостающей») комплементарной ей цепи за счет свободных нуклеотидов, всегда имеющихся в большом количестве в клетке. В результате вместо одной («материнской») молекулы ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует клеточному делению и обеспечивает передачу наследственной информации от материнской клетки дочерним и всем последующим.
14 . Рибонуклеиновые кислоты, их виды, строение, назначение.
РНК - класс нуклеиновых кислот,линейных полимеровнуклеотидов, в состав которых входят остаток фосфорной кислоты, рибоза (в отличие отДНК, содержащей дезоксирибозу) и азотистые основания -аденин,цитозин,гуанини урацил (в отличие от ДНК, содержащий вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусов. РНК содержатся главным образом вцитоплазме клеток. Эти молекулы синтезируются в клетках всех клеточных живых организмов, а также содержатся в вироидах и некоторых вирусах. Основные функции РНК в клеточных организмах - это шаблон для трансляции генетической информации в белки и поставка соответствующих аминокислот к рибосомам. В вирусах является носителем генетической информации (кодирует белки оболочки и ферменты вирусов). Структура РНК .
Молекула имеет однонитевое строение. Полимер. В результате взаимодействия нуклеотидов друг с другом молекула РНК приобретает вторичную структуру, различной формы (спираль, глобула и т.д.). Мономером РНК является нуклеотид (молекула, в состав которой входит азотистое основание, остаток фосфорной кислоты и сахар (пептоза)). РНК напоминает по своему строению одну цепь ДНК. Нуклеотиды, входящие в состав РНК: гуанин, аденин, цитозин, урацил. Аденин и гуанин относятся к пуриновым основаниям, цитозин и урацил к пиримидиновым. В отличие от молекулы ДНК, в качестве углеводного компонента рибонуклеиновой кислоты выступает не дезоксирибоза, а рибоза. Вторым существенным отличием в химическом строении РНК от ДНК является отсутствие в молекуле рибонуклеиновой кислоты такого нуклеотида как тимин. В РНК он заменён на урацил.
Виды и типы РНК клеток.
Существуют три типа РНК, каждый из которых выполняет свою особую роль в синтезе белка.
1. Матричная РНК переносит генетический код из ядра в цитоплазму, определяя таким образом синтез разнообразных белков.
2. Транспортная РНК переносит активированные аминокислоты к рибосомам для синтеза полипептидных молекул.
3. Рибосомная РНК в комплексе примерно с 75 разными белками формирует рибосомы - клеточные органеллы, на которых происходит сборка полипептидных молекул.
Матричная РНК представляет собой длинную одноцепочечную молекулу, присутствующую в цитоплазме. Эта молекула РНК содержит от нескольких сотен до нескольких тысяч нуклеотидов РНК, образующих кодоны, строго комплементарные триплетам ДНК.
Еще один тип РНК, играющий важнейшую роль в синтезе белка, называют транспортной РНК , поскольку он транспортирует аминокислоты к строящейся молекуле белка. Каждая транспортная РНК специфически связывается только с одной из 20 аминокислот, составляющих белковые молекулы. Транспортные РНК действуют как переносчики специфических аминокислот, доставляя их к рибосомам, на которых происходит сборка полипептидных молекул.
Каждая специфическая транспортная РНК распознает «свой» кодон матричной РНК, прикрепившейся к рибосоме, и доставляет соответствующую аминокислоту на соответствующую позицию в синтезируемой полипептидной цепи. Цепь транспортной РНК гораздо короче матричной РНК, содержит всего около 80 нуклеотидов и упакована в форме клеверного листа. На одном конце транспортной РНК всегда находится аденозинмонофосфат (АМФ), к которому через гидроксильную группу рибозы прикрепляется транспортируемая аминокислота. Транспортные РНК служат для прикрепления специфических аминокислот к строящейся полипептидной молекуле, поэтому необходимо, чтобы каждая транспортная РНК обладала специфичностью и в отношении соответствующих кодонов матричной РНК. Код, посредством которого транспортная РНК распознает соответствующий кодон на матричной РНК, также является триплетом и его называют антикодоном. Антикодон располагается примерно посередине молекулы транспортной РНК. Во время синтеза белка азотистые основания антикодона транспортной РНК прикрепляются с помощью водородных связей к азотистым основаниям кодона матричной РНК. Таким образом, на матричной РНК выстраиваются в определенном порядке одна за другой различные аминокислоты, формируя соответствующую аминокислотную последовательность синтезируемого белка.
Макромолекулярная структура ДНК
Выделение дезоксирибонуклеиновых кислот
Выделение рибонуклеиновых кислот
Природа межнуклеотидных связей
Межнуклеотидная связь в ДНК
Межнуклеотидная связь в РНК
Значение нуклеиновых кислот
Список литературы
1. Состав нуклеиновых кислот
Нуклеиновые кислоты - это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев - нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида - структурного звена нуклеиновых кислот - входят три составные части:
азотистое основание - пиримидиновое или пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
моносахарид - рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот – рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота.
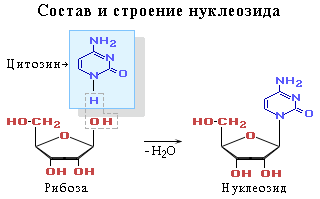
Нуклеотид - фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
В конце 40-х - начале 50-х годов, когда появились такие методы исследования, как хроматография на бумаге и УФ-спектроскопия, были проведены многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Полученные данные позволили решительно отбросить старые представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности (так называемая тстрануклеотидная теория строения ПК. господствовавшая в 30-40-е годы), и подготовили почву для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях.
Метод определения состава ПК основан на анализе гндролизатов, образующихся при их ферментативном или химическом расщеплении. Обычно используются три способа химического расщепления НК. Кислотный гидролиз в жестких условиях (70%-ная хлорная кислота, 100°С, 1 ч или 100%-ная муравьиная кислота, 175 °C, 2 ч), применяемый для анализа как ДНК, так и РНК, приводит к разрыву всех N-гликозидных связей и образованию смеси пуриновых и пиримидиновых оснований. При исследовании РНК могут использоваться как мягкий кислотный гидролиз (1 н. соляная кислота, lOO°C, 1 ч), в результате которого образуются пуриновые основания и пирамидиповые нуклеозид-2"(3")-фосфаты, так и щелочной гидролиз (0,3 н. едкий кали, 37 °С, 20 ч), дающий смесь нуклеозид -2" (3") -фосфатов.
Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК.
При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки).
Использование описанных выше аналитических приемов показало, что ПК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах.


